Compound or attached-X chromosomes (denoted X^X) consist of two full-length X chromosomes sharing a common centromere so that they are always inherited together.
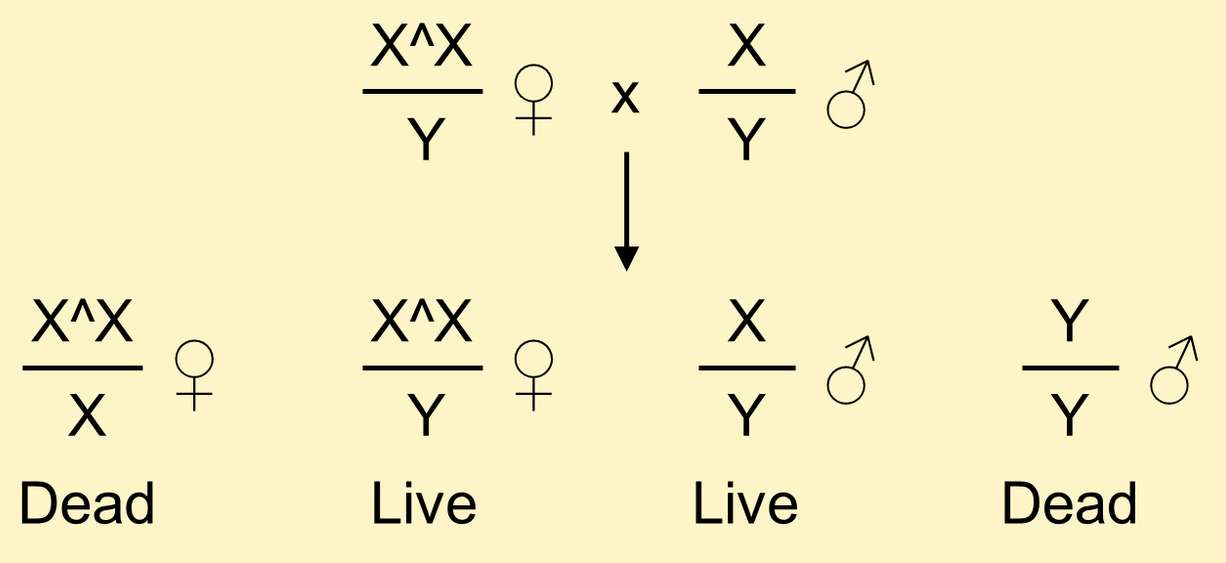
In crosses of normal males to females carrying an attached X, male progeny inherit their X from their father and their Y from their mother.
Stable stocks of X-linked visible and female-sterile mutations can be established "instantly" by crossing mutant males to females carrying an attached X.
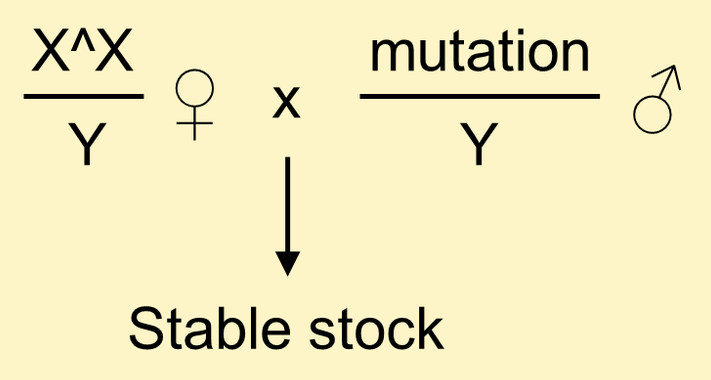
Maintaining mutations in this way prevents spontaneous lethal and male-sterile mutations from accumulating on the X chromosomes.
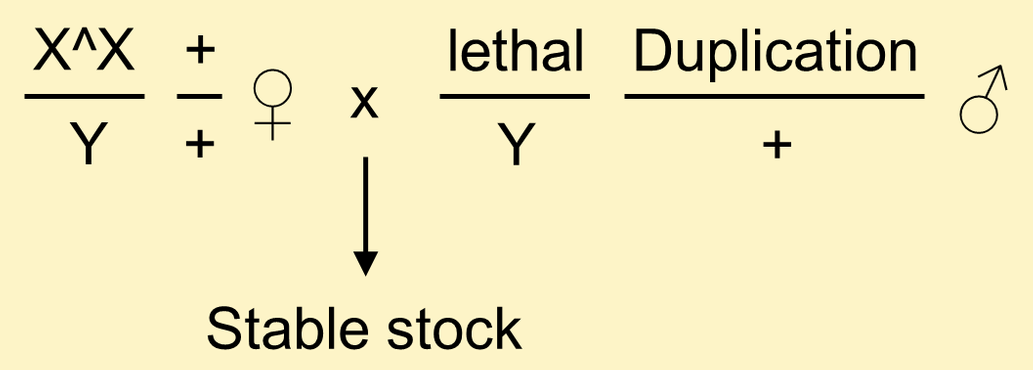
It is possible to maintain X-linked lethal mutations in stock without accumulating additional lethals on the chromosome if you include a rescuing transgene or chromosomal duplication.
Attached-X stocks are also useful for maintaining attached-XY chromosomes in the absence of a free Y chromosome.
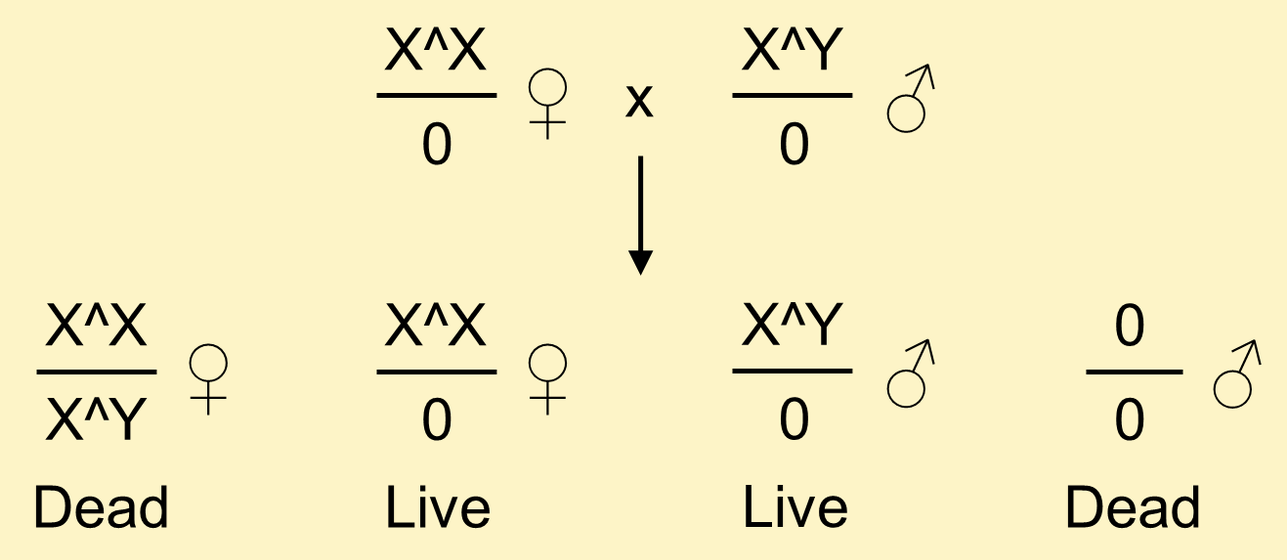
The simplest compound-X chromosome is the "reversed metacentric", denoted C(1)RM, which is formed by a reciprocal translocation with one break in centric heterochromatin to the left of the centromere and the other break in heterochromatin to the right of the centromere.
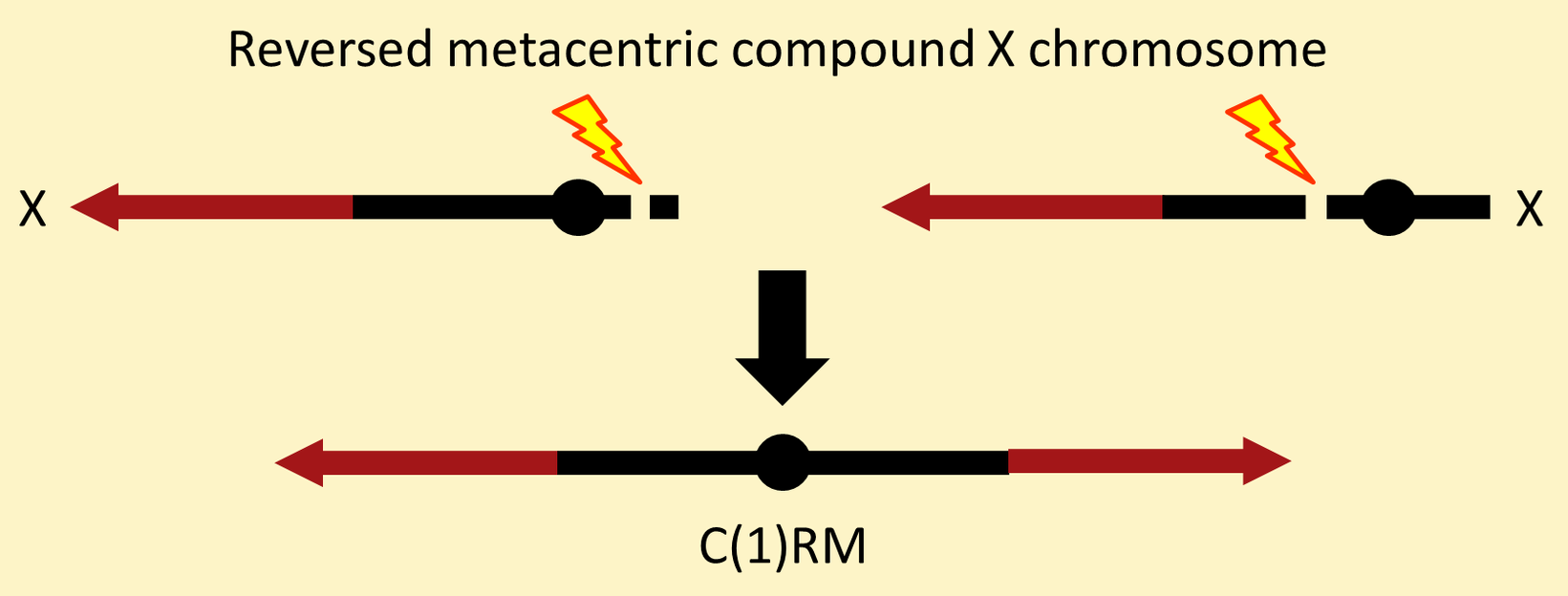
It is called a reversed metacentric because the centromere is in the middle (metacentric) and the two component X chromosomes lie in opposite orientations.
C(1)RM chromosomes are available with many different marker mutations on the arms. The markers are usually homozygous, because meiotic crossovers between the two component X chromosomes lead to loss of heterozygosity within a few generations. Here are the marker combinations available in Bloomington stocks.
y[1] w[1] f[1] | y[1] v[1] f[1] mal[1] | y[2] su(w[a])[1] w[a] | y[1] | y[1] shi[1] f[1] | y[1] v[1] f[1] |
y[1] v[1] | y[1] v[1] f[1] mal[2] | unmarked | y[1] pn[1] v[1] | y[1] bb[*] | sc[1] v[1] f[1] |
y[1] v[1] bb[1] | y[2] | ras[l1] | y[1] pn[1] | y[1] B[1] | |
Fortunately, you don't need to induce chromosome breaks to generate a new C(1)RM. Instead, you can use a "reversed metacentrigenic" chromosome. An "RMG" chromosome is a full X with the base of another X appended as a right arm. Meiotic recombination between a normal X and the short right arm generates a new C(1)RM.
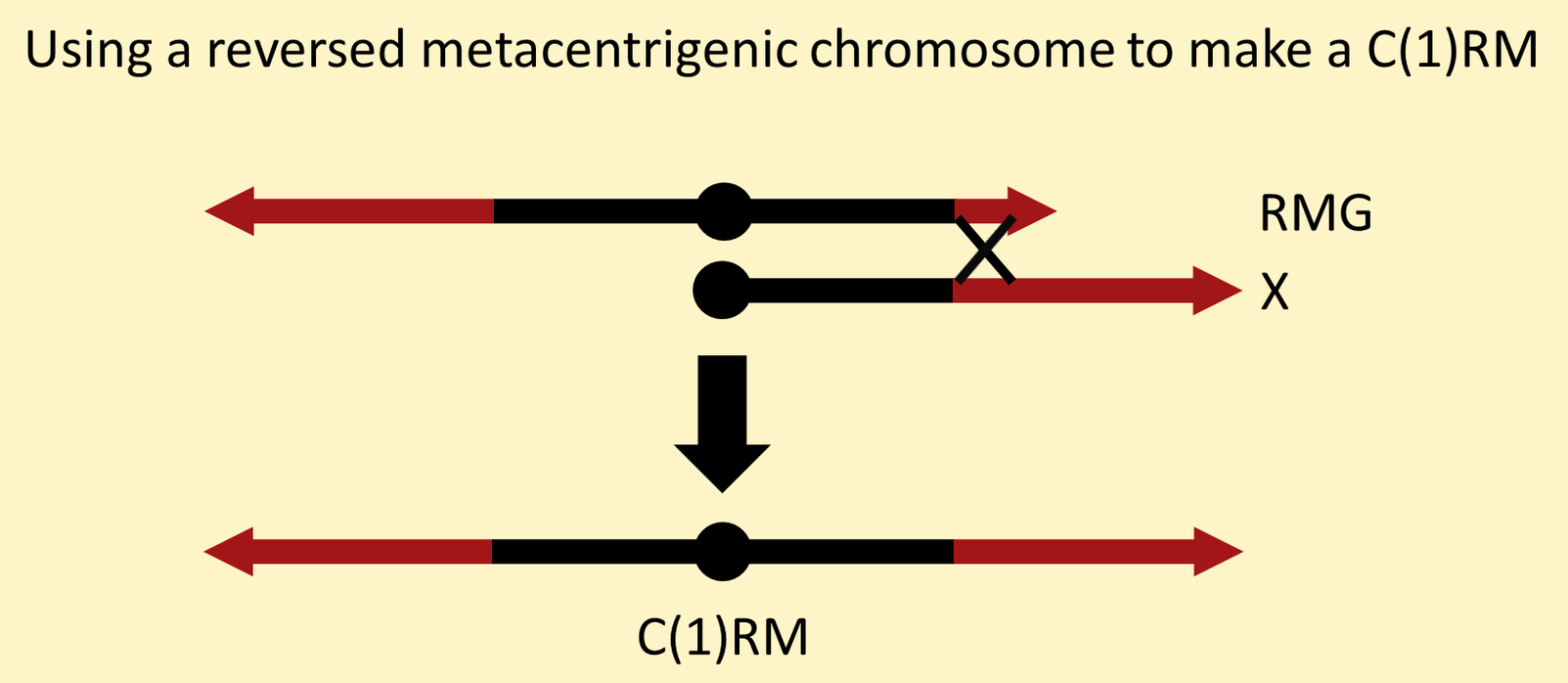
We have an RMG chromosome called Dp(1;1)MMG that can be used to make an C(1)RM with any combination of alleles you care to use.
| Stock # | Genotype |
|---|
| 6467 | Dp(1;1)MMG, y[2] cv[1] v[1] f[1], B[S]; ins[1] |
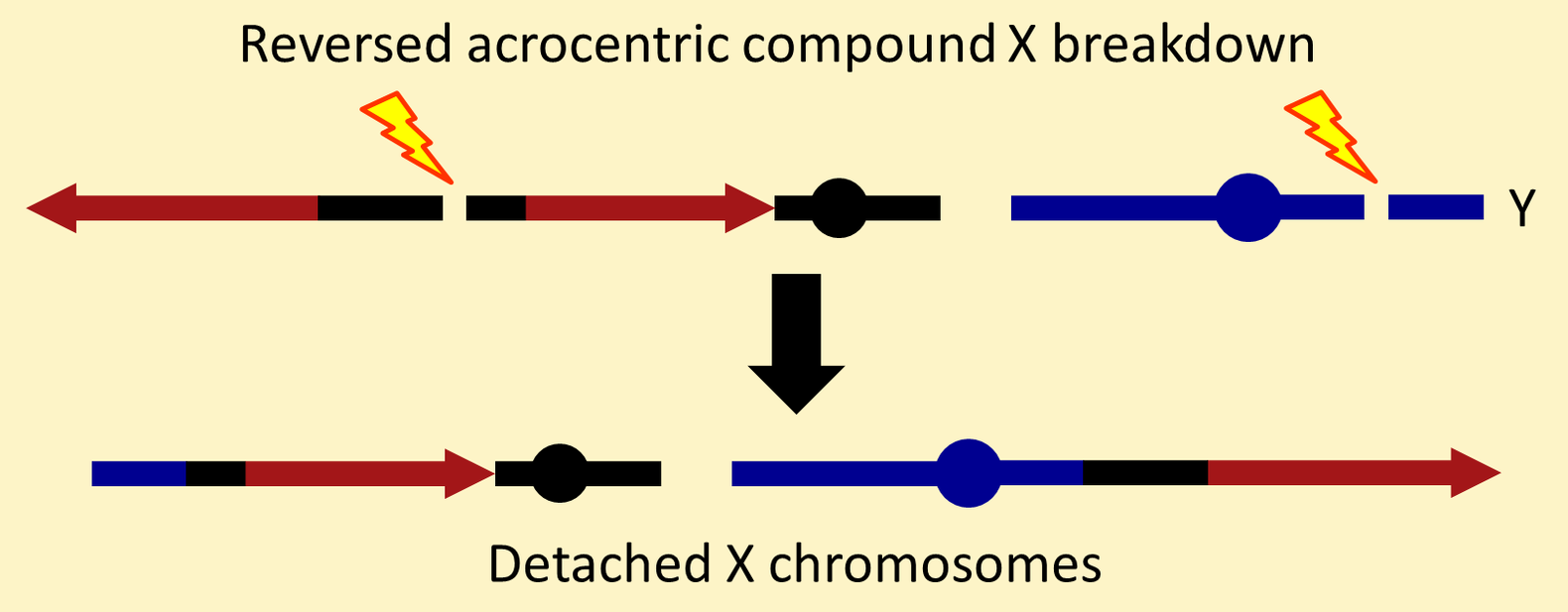
Unfortunately, C(1)RM chromosomes break down at a high frequency from spontaneous DNA breaks in heterochromatic regions. Usually, the component X chromosomes "detach" following reciprocal translocation between X centric heterochromatin and Y heterochromatin.
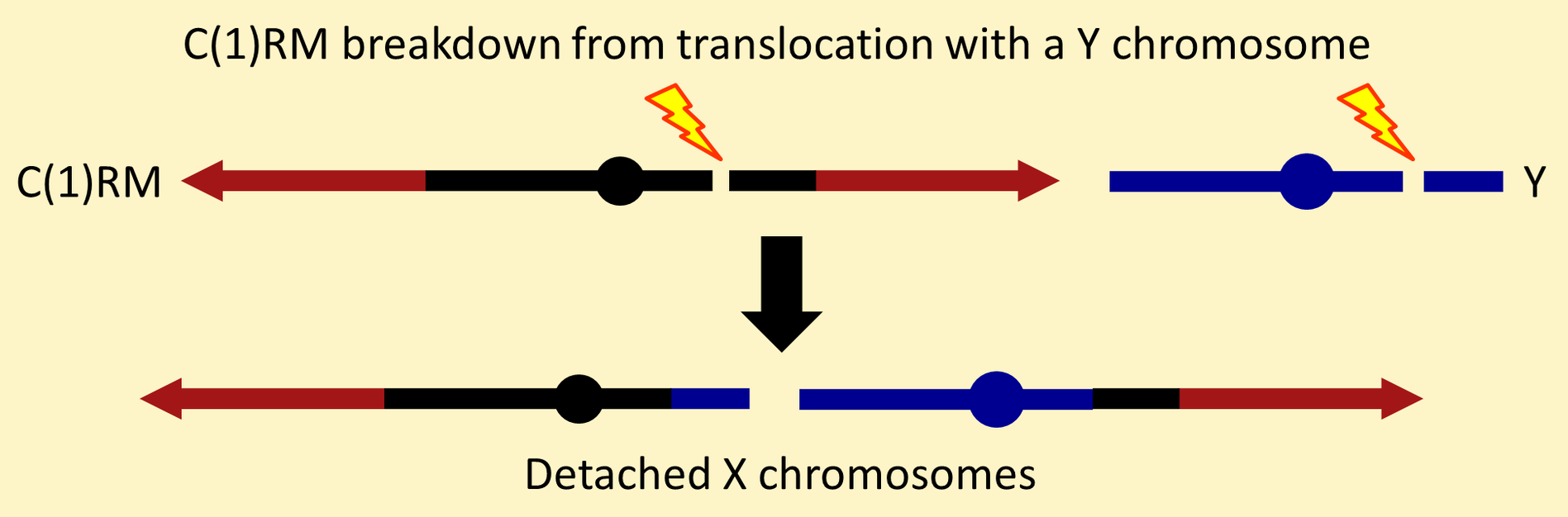
This is the reason that C(1)RM chromosomes are more stable if they are maintained in stocks in the absence of a Y chromosome. Attached-XY chromosomes allow you to construct stocks where C(1)RM is never combined with a Y in females.
Most of the other attached-X chromosomes in common use are "reversed acrocentric" chromosomes. They start with an inversion that encompasses almost all of an X chromosome.
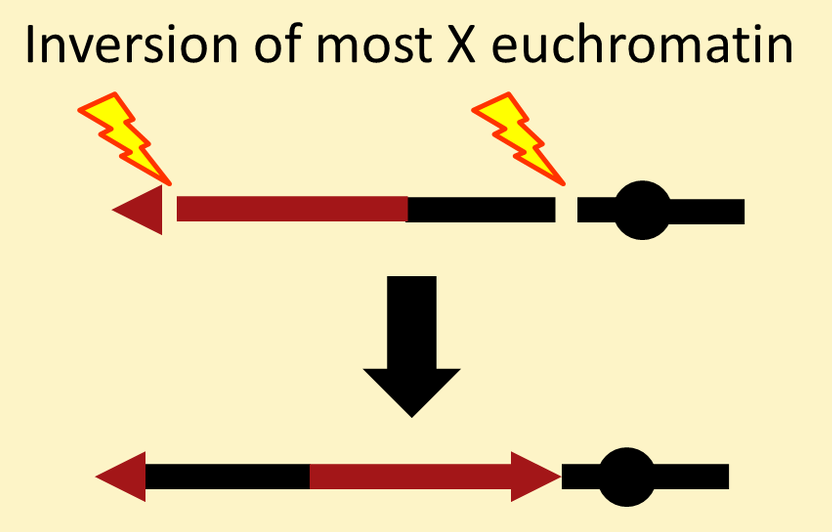
Then a reciprocal translocation between the base of another X and the distal block of heterochromatin on the inversion chromosome creates an attached-X with the centromere at one end (acrocentric) and the two component X chromosomes lying mostly in opposite ("reversed") orientations.
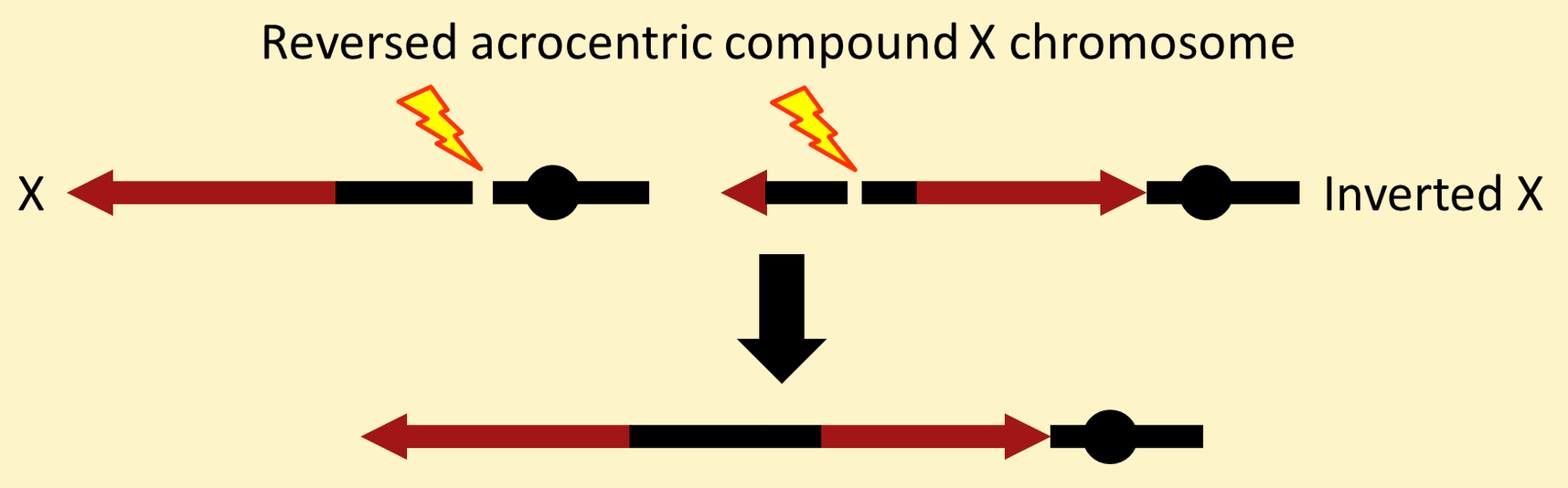
The reversed acrocentric chromosome with this simple structure is called C(1)RA. It is available with the following markers.
| y[1] | cin[1] y[1] | v[1] f[1] | y[1] l(1)1Ac[1] |
The other reversed acrocentric chromosomes in Bloomington stocks—C(1)DX, C(1)M3 and C(1)M4—have additional inverted internal segments.
Unfortunately, reversed acrocentric chromosomes also break down from spontaneous heterochromatic breaks—especially breaks in the medial heterochromatic block and in a Y chromosome.
C(1)DX is popular because it is more stable than most attached-X chromosomes. The reason is that the medial heterochromatic block in C(1)DX is particularly small and, consequently, it is a smaller target for translocation events leading to detachment.
All C(1)DX chromosomes are marked with y[1] and f[1]. Some carry mutations in the white gene, too.
A drawback to C(1)DX is that the reciprocal translocation event that attached the X chromosomes eliminated the tandem arrays of ribosomal RNA genes (the bobbed locus) from the heterochromatin of both component X chromosomes. Consequently, females with C(1)DX must carry a Y chromosome providing the Y-linked rRNA genes to live. For most simple uses of C(1)DX, this does not matter.
C(1)M3 and C(1)M4 combine a balancer X chromosome carrying the centromere with another distally located X chromosome with internal inversions. Both chromosomes are available with y[1] or y[2] markers.
C(1)M4 is interesting because one of the internal inversions in the distal component X chromosome, In(1)w[m4], causes position effect variegation of the w gene. Variegation is strong in the absence of a Y chromosome, but it is suppressed when a Y chromosome is present.
The medial block of heterochromatin in C(1)M3 and C(1)M4 is larger than the block present in C(1)DX, so detachment is more common. Roughly 1 in 15,000 progeny of C(1)M4/Y females carries a detached X from reciprocal translocation between C(1)M4 and the Y.
C(1)M5 is present in a couple of Bloomington stocks. It is probably another reversed acrocentric, but we do not know its origin. It seems relatively stable. Some stocks with C(1)M5 show variegated eye pigmentation, suggesting that C(1)M5 carries In(1)w[m4] like C(1)M4.
The final attached-X chromosome present in Bloomington stocks is C(1)A. Its history is considerably more complicated than the other attached-X chromosomes (see Traverse & Pardue (1988)). Its component segments are arranged in the order
13E — 7A1 | 7A1 — 20.1 — 6F2 | 6F2 — 1 | 20 — 13E
It is marked with y[1] and it may be used in stocks like any other attached-X. Despite claims that it is the most stable attached-X, it breaks down at a rate similar to most other attached-X chromosomes—though the mechanism of breakdown has not been explored.
Conclusion
Females carrying any of these attached-X chromosomes are relatively healthy and fertile, so your choice of chromosome will likely come down to markers, the need for a free Y and the risk of detachment. All of these attached-X chromosomes will work for most stockkeeping purposes as long as you monitor your stocks periodically for breakdown.
Reference
For a more detailed discussion of compound X chromosomes, see Chapter 19 of Ashburner, Golic and Hawley (2005) “Drosophila: a Laboratory Handbook”.
Miscellaneous information
C(1)DX was named “Double X”. Originally, it was denoted := and this symbol can be seen in some older papers.
“Attached-X” was originally used only for C(1)RM even though it is now used generically for all compound-X chromosomes. C(1)RM is denoted ·= in older papers. It was the first compound chromosome isolated. The amusing story of its discovery is copied here.
We have chromosomes similar to Dp(1;1)MMG, the reversed metacentrigenic chromosome described above, for generating your own tandem acrocentric and tandem metacentric compound-X chromosomes. They are TMG (tandem metacentrigenic) and TAG (tandem acrocentrigenic).
| Stock # | Genotype |
|---|
| 981 | Dp(1;1)B[S]TMG, y[1] sc[4] m[1] f[1] B[S]/In(1)sc[7], In(1)AM, sc[7] ptg[4] v[1] |
| 4484 | Dp(1;1)B[S]TAG, sc[1] w[1] B[BB]/C(1)DX, y[1] f[1] |
Stocks
For a curated list of attached-X stocks with examples of each marker variant see here.
For a complete list of attached-X stocks in the Bloomington collection see here.

