

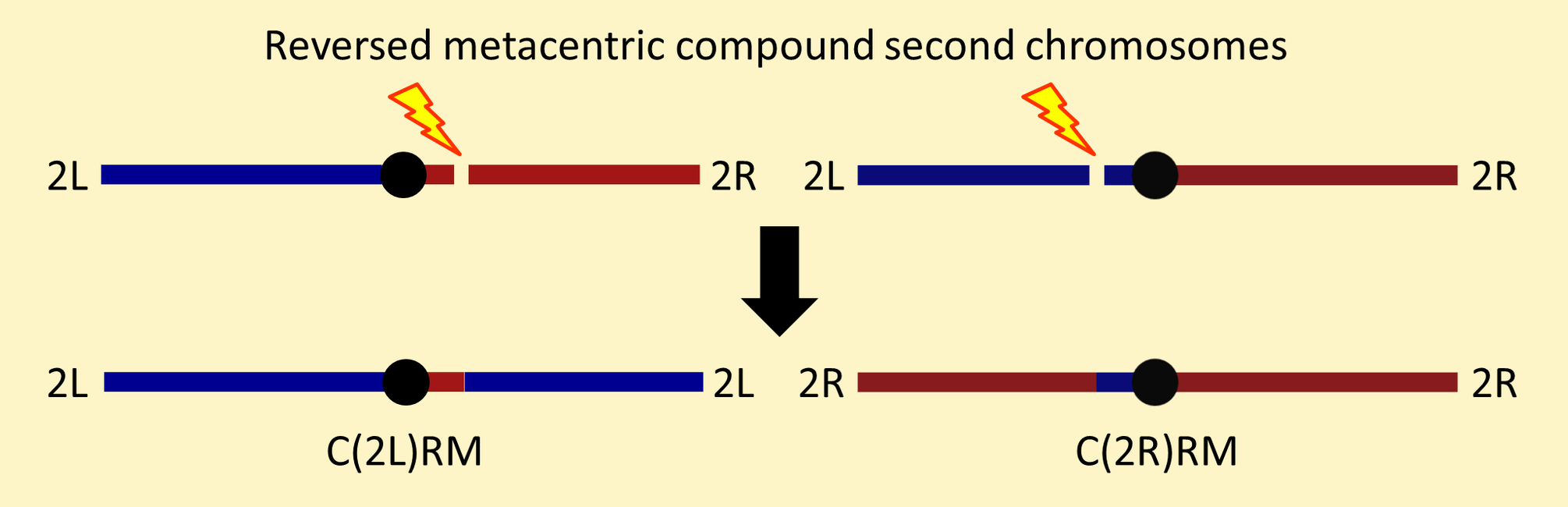
In compound chromosomes, homologous chromosome arms are attached to the same centromere. The compound autosomes in the Bloomington collection come in two forms as shown in the following figures.
Reversed metacentric chromosomes arise from reciprocal translocation events involving breaks in centric heterochromatin on opposite arms as shown below.
C(2)EN was constructed by appending Y chromosome segments to the ends of C(2L)RM and then appending 2R arms to those chromosome ends. C(3)EN was constructed similarly.
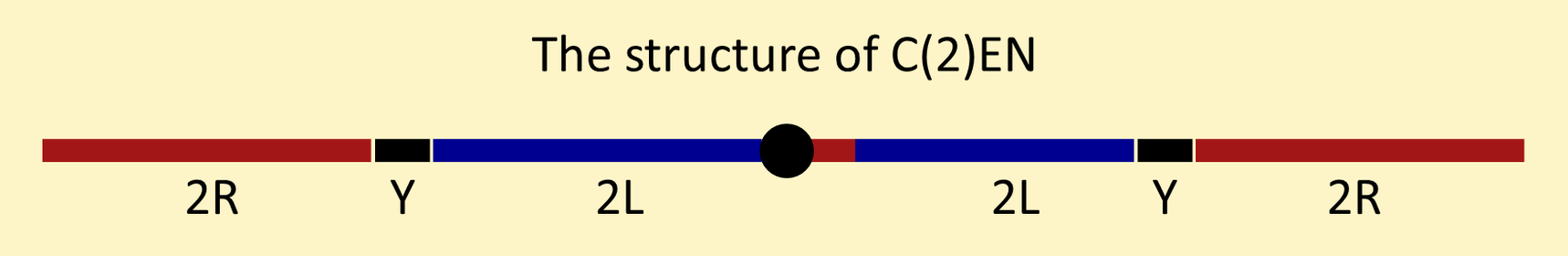
Males carrying C(2L)RM and C(2R)RM make four kinds of sperm in equal proportions: C(2L)RM + C(2R)RM sperm, nullo-2 sperm, C(2L)RM-only sperm and C(2R)RM-only sperm. Males carrying C(2)EN make only C(2)EN and nullo-2 sperm.
Compound autosomes are used most often to detect primary nondisjunction in females. For example, diplo-2 eggs arising from nondisjunction will give rise to euploid zygotes if they are fertilized by nullo-2 sperm produced by males carrying compound autosomes and vice versa. Normal eggs produce aneuploid, lethal zygotes in these crosses.
Compound fourth chromosomes are also used to detect nondisjunction, but mono-4 and triplo-4 zygotes are viable.
Good overviews of compound autosomes are given in chapter 19 of Drosophila: a laboratory handbook and in Holm (1976).
The recessive marker mutations in the following stocks are present on both arms of the compound chromosomes.